Course Authors
Juan F. López, M.D.
Release Date: 05/08/2002
Upon completion of this Cyberounds®, you should be able to:
List some of the signs and symptoms of Major Depressive Disorder
Describe research findings showing hyperactivity of the Hypothalamic-Pituitary Adrenal Axis in Major Depressive Disorder
Describe some of the serotonin receptor changes found in the brains of suicide victims
Discuss some of the ways in which the Hypothalmic-Pituitary-Adrenal Axis and the Serotonin system interact
Discuss the evidence linking stress with the onset of depressive symptoms.
Introduction
Major Depressive Disorder (MDD), also known as Major Depression, is a psychiatric syndrome characterized by pervasive disturbances in mood, sleep, appetite, energy, motivation, hedonic capacity and thinking. According to the Diagnostic and Statistical Manual of Mental Disorders (DSM-IV), a depressive episode must be diagnosed if a patient has had depressed mood, or has lost interest or pleasure in most activities, for a duration of at least two weeks. However, depressive episodes often last months, sometimes years, and they carry a significant impairment in social and occupational functioning. Depressive episodes also tend to be recurrent and, if left untreated, most patients will have multiple episodes during their lifetime. The episodes tend to become more frequent and/or more severe as the disease progresses.(61)
Table 1. Diagnostic criteria for a Major Depressive Episode (Adapted from DSM-IV).
At least five of the following symptoms have been present during the same two-week period and represent a change from previous functioning:
|
MDD is a common medical disorder, with a lifetime prevalence of 17.1% and a 12-month prevalence of 10.3% in the general population.(36) However, the prevalence of MDD in medical populations is even higher, with rates ranging from 30% to 50%, depending on the specific medical condition. This psychiatric disorder also carries a significant morbidity and mortality with a negative impact not only for the patient itself but for the family and for society in general. About two thirds of severely depressed patients exhibit suicidal ideation and 10% to 15% commit suicide.(31)
The specific pathogenetic cause of depression is unknown but it is widely accepted that its etiology, course and long-term prognosis are influenced by genetic, environmental and neurobiological factors. A greater understanding of the neurobiology of this disorder is leading us to better treatment strategies that can prevent or decrease the impact of this disease. Although many neurotransmitters and neurohormones have been linked to the pathophysiolgy of depression (e.g., norepinephrine, dopamine, thyroid hormones), research studies have implicated disturbances in the serotonin (5-HT) system and the Hypothalamic-Pituitary-Adrenal (HPA) axis as two of the neurobiological alterations most consistently associated with affective illness.(25),(52),(33),(57)
Historically, the role of each of these two systems in mood disorders has been studied independently, but more recently their interaction in the brain, as it relates to the pathophysiology of depression, has received increased attention. In this Cyberounds®, we will review recent animal and human findings, from the perspective of the 5-HT and HPA axis, which shed new light on the neurobiology of MDD. We will discuss some of the ways in which adrenal glucocorticoids and 5-HT receptors interact during conditions of chronic stress or severe "allostatic load. We will also discuss what is the potential significance of these findings for the pathophysiology and treatment of mood disorders.
That disturbances in the Hypothalamic-Pituitary-Adrenal axis and in serotonin may share a common pathophysiological mechanism is not surprising, since we know from animal studies that they interact extensively and that they are related in a variety of ways.(49),(11),(46) The hippocampus and the paraventricular nucleus of the hypothalamus (PVN), in particular, are anatomical regions in which components of the Hypothalamic-Pituitary-Adrenal axis and the 5-HT systems have a rich representation (see Figure 1 for an illustration of how serotonin neurons from the raphe nucleus project to the PVN and hippocampus). These regions are also part of the limbic system, an area implicated in the regulation of several vegetative functions (arousal, sleep, appetite and hedonic capacity) as well as in the control of mood. The hippocampus has also been implicated in memory and cognitive function. The recognition that the hippocampus is an integral component of the Hypothalamic-Pituitary-Adrenal axis has led some investigators (including our laboratory) to refer to this neuroendocrine system as the "Limbic"-Hypothalamic-Pituitary-Adrenal (LHPA) axis. We will, therefore, for the purpose of this presentation, use the term "LHPA", instead of the more commonly used term "HPA", when referring to this system.
Figure 1. Serotonin Neurons and the LHPA Axis.
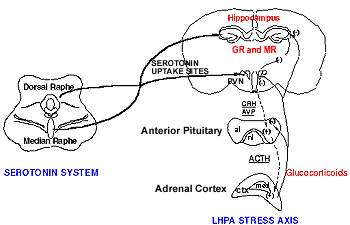
Studies have shown that the Serotonin (5-HT) system and the Limbic Hypothalamic Pituitary-Adrenal (LHPA) axis are linked in a variety of ways.
| GR = glucocorticoid receptor MR = mineralocorticoid receptor PVN = Paraventricular nucleus of the hypothalamus CRH = corticotropin releasing hormone AVP = vasopressin ACTH = adrenocorticotropin hormone |
al = anterior lobe of the pituitary nl = neural lobe ctx = adrenal cortex med = medulla (+) denotes stimulation (-) denotes inhibition. |
The Limbic-Hypothalamic-Pituitary-Adrenal Axis and Stress
As discussed elsewhere in this series, the LHPA is the classic neuroendocrine system that responds to stress. Perception of stress by an organism results in a series of events, the final result of which is the secretion of glucocorticoids (cortisol in humans, corticosterone in rats) from the adrenal cortex.(16) Activation and termination of the adrenocortical stress response is critical for adaptation and survival. Inhibition of stress responsiveness is partly achieved by the binding of circulating glucocorticoids to specific cytoplasmic receptors in hypothalamus, where they inhibit corticotropin releasing hormone (CRH) and, consequently, pituitary adrenocorticotropin (ACTH) secretion. Additional modulation of the system is apparently achieved in limbic structures, especially the hippocampus, a structure that is linked to the hypothalamus through neuronal connections that converge on the paraventricular nucleus of the hypothalamus (PVN), where the stress responsive CRH and vasopressin (AVP) neurons reside.(42)
There are several lines of evidence that highlight the importance of the hippocampus for LHPA feedback mechanisms.(50) Pioneer work by McEwen demonstrated that the hippocampus contains a high abundance of two types of glucocorticoid receptors which are thought to control negative feedback: Type I (also known as Mineralocorticoid Receptor, or "MR") and Type II (also known as Glucocorticoid receptors or "GR"). Type I (or MR) resembles the kidney mineralocorticoid receptor and has stringent specificity, binding selectively corticosterone, the main glucocorticoid of the rat. In the rodent brain, MR is most densely localized in hippocampal and septal neurons. Type II or GR receptors are widely distributed in rat brain, including hippocampus, hypothalamus and prefrontal cortex. However, they bind corticosterone with a lower affinity compared to MR. Their highest affinity is for potent synthetic glucocorticoids, such as dexamethasone.(50) These receptor characteristics complement each other and put the MR and GR in a position to modulate LHPA responses (see DeKloet et al.(20) for a recent excellent review on the neurobiology of GR and MR). The MR receptors appear to be operative at low corticosterone concentrations and may offer tonic inhibition to the axis during the nadir of the circadian rhythm.(19),(50) When high concentrations are present, MR receptors saturate and the GR receptors ensure the return of homeostasis. As stated above, it is also well established that the hippocampus is a central component of limbic circuitry and is fundamental in controlling aspects of cognitive and behavioral functions, putting these receptors in a position to modulate, simultaneously, the neuroendocrine and cognitive response of the organism to stress.
The Limbic-Hypothalamic-Pituitary-Adrenal Axis in Depression
Hyperactivity of the LHPA axis is a well documented phenomena in MDD. This dysregulation is manifested, among other things, by cortisol hypersecretion, failure to suppress cortisol secretion after dexamethasone administration, exaggerated adrenal responses to endocrine challenges and blunted ACTH response to CRH administration.(9),(30),(41),(25),(33) This last observation has been interpreted as evidence of pituitary downregulation of CRH receptors, secondary to an increase in secretion of CRH. There is indeed good evidence of increased central drive, based on increased activity at the nadir of the circadian rhythm,(73) as well as more direct findings of elevated CRH in the CSF of depressed patients(55) and increased CRH immunoreactivity and mRNA levels in the PVN.(62),(63) Interestingly, post-mortem studies have also found evidence of chronic LHPA activation in suicide victims, such as adrenal hyperplasia,(22) downregulation of CRH receptors(56) and increases in proopiomelanocortin mRNA, the precursor for ACTH, in the pituitary.(43) It is not known whether these LHPA changes in suicide are due to the fact that a significant subset of suicide victims are patients with depressive disorders, to the stress surrounding the suicide itself or to a neurobiological "abnormality" common to all suicides irrespective of diagnosis.
We have found, in a group of suicide victims with a history of depression, decreases in hippocampal MR mRNA levels(45) and, more recently, a decrease in GR mRNA levels in prefrontal cortex (unpublished observation), an area of the brain which is associated with higher cognitive and executive function. We don't know whether these changes represent a genetic or developmental, vulnerability "marker" for suicide or for depression. Nevertheless, these findings are consistent with a history of exposure to chronic stress and/or to high peripheral glucocorticoid levels.(27),(28)
Historically, the presence of LHPA overactivity in patients with depression was believed by many to be a "secondary" phenomena of the illness, reflecting either a central monoaminergic disturbance, the stress of the illness or both. However, over the past few years, it has become clearer that the LHPA abnormalities in MDD are intimately linked to the pathohysiology of the disease. This change in perspective was stimulated, in part, by the increased awareness that glucocorticoids, the final products of the LHPA axis, have been shown to have profound effects on mood and behavior.(49) For example, a high incidence of depression is linked to pathologies involving elevated corticosteroid levels, such as Cushing's syndrome. This corticosteroid-induced depression usually disappears when corticosteroid levels return to normal.(32),(54) In fact, it has become increasingly clear, from both animal and clinical studies, that circulating glucocorticoid levels provide important hormonal control of affect, which may be mediated by steroid-induced modulation of central limbic circuitry.(49) The precise mechanism by which corticosteroids exert this influence on affect is not well understood. However, this mechanism is likely to involve interactions with brain neurotransmitters, since we know that central control of affect is intimately associated with the actions of the monoamines serotonin, norepinephrine and dopamine.
Serotonin Receptors and Depression
The serotonin system has been widely investigated as a key element in the pathophysiology of depression(52) and of suicide,(48) and as a mediator of the therapeutic action of antidepressants.(3) Although the 5-HT system has many components, the three 5-HT molecules believed to be most closely associated with the neurobiology of mood are the serotonin transporter (5-HTt), the serotonin 1a receptor (5-HT1a) and the serotonin 2a receptor (5-HT2a, formerly 5-HT2). Most newer (as well as older) antidepressants inhibit the re-uptake of serotonin from the synapse and alter 5-HTt protein and mRNA levels.(44),(58) Animal studies have also demonstrated that chronic antidepressant administration affects the function and number of the 5-HT1a and 2a receptors. Many electrophysiological studies have shown that antidepressants "upregulate" or "sensitize" 5-HT1a function in the hippocampus, while at the same time they "down-regulate" or "desensitize" 5-HT1a function in the raphe, where it acts as an inhibitory somatodendritic receptor.(5) Some studies have also reported a modest increase in 5-HT1a receptor number in hippocampus following antidepressant administration to rodents.(70),(37) We have found that suicide victims, with a history of depression, have decreases in 5-HT1a gene expression in hippocampus.(45) These changes are in the opposite direction of what is observed with antidepressant treatment in rat hippocampus.
The 5-HT2a receptor has also been shown to be affected by chronic antidepressant treatment. Most, but not all, studies have reported decreases in 5-HT2a binding in the prefrontal cortex after chronic antidepressant administration.(60),(7) The opposite changes (5-HT2a upregulation) are found in the prefrontal cortex of suicide victims,(1),(29) although these findings are not universal.(68) In addition, subjects with a history of MDD dying of natural causes have increases in 5-HT2a binding in the prefrontal cortex.(72)
These findings have led several investigators to propose that postsynaptic 5-HT1a and 5-HT2a receptors have functionally opposing effects, that a disturbed balance of these receptors may be contributing to the pathophysiology of depression(3) and that restoring this balance is necessary for antidepressant action.(6)
Interaction Between the LHPA Axis and Serotonin
Another level of complexity is added by the fact that 5-HT and the LHPA axis interact at multiple levels. For example, it is known that some of the 5-HT neurons arising from the nucleus raphe dorsalis and nucleus raphe magnus project to the PVN and synapse onto CRH neurons.(24) 5-HT neurons also project to other brain areas, such as the amygdala and the uprachiasmatic nucleus, which are thought to modulate the function of the PVN.(69) Pharmacological stimulation with 5-HT agents can activate ACTH and cortisol release.(24) Many brain areas, which express 5-HT receptors, also have abundant concentrations of corticosteroid receptors. In the limbic system in particular, the hippocampus has high concentrations of 5-HT1a in the same subfields that contain abundant GR and MR receptor levels (Figure 2). The prefrontal cortex is rich in 5-HT2a receptors,(59) as well as GR receptors.
Figure 2.
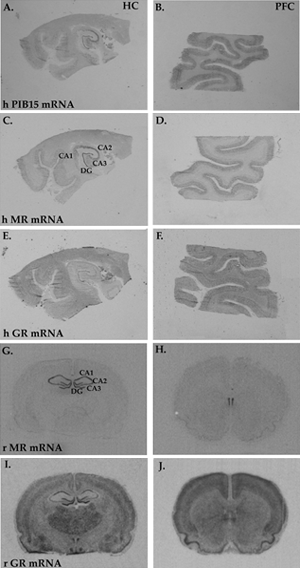
Figure 2 shows photomicrographs comparing the distribution of different mRNAs in human and rat brain, by in situ hybridization. The distribution of P1B15 mRNA, a gene encoding for the molecule cyclophillin is shown in plates A and B, for comparison purposes. P1B15 is often used in postmortem studies to correct for mRNA degradation. MR mRNA (C,D) and GR mRNA (E,F) are shown in adjacent sections of human hippocampus (left column) and prefrontal cortex (right column). Equivalent regions in rat brain are shown, comparing the distribution of MR mRNA (G,H) and GR mRNA (I,J). Notice the widespread distribution of GR in brain. Notice also the low abundance of MR mRNA in rat cortex compared to human.
HC = hippocampus, PFC = prefrontal cortex, h = human, r = rat.
Interestingly, unlike rodents, human prefrontal cortex also has significant amounts of MR mRNA (Figure 2). This constitutes anatomical evidence suggesting that MR may have a more expanded role in modulating brain function in humans than in rodents. Furthermore, the co-localization of MR and GR in prefrontal cortex suggest that both of these receptors are capable of modulating higher brain functions, such as mood, social behavior and cognitive processing, perhaps by interacting with 5-HT receptors.
Serotonin and corticosteroid receptors not only interact anatomically but also functionally. It has been reported that administration of serotonin can up-regulate GR in the hippocampus and that, conversely, pharmacological destruction of serotonergic projections decreases GR and MR mRNA levels in the hippocampus.(4),(66) Regulation in the other direction, i.e., glucocorticoid regulation of 5-HT receptors, has also been reported. Animal studies have demonstrated that adrenalectomy and corticosteroid administration powerfully regulate 5-HT1a receptor number and mRNA in the hippocampus.(10),(12),(40) This effect of glucocorticoids seems to be specific for the postsynaptic 5-HT1a receptor, since the somatodendritic 5-HT1a is not affected by these manipulations.(10)
The 5-HT2a receptor is also sensitive to changes in peripheral glucocorticoid levels. Upregulation of 5-HT2a in rat neocortex cortex has been reported following ten days of exogenous administration of ACTH.(38) This effect is abolished by adrenalectomy and mimicked by corticosterone administration for ten days. Dexamethasone treatment for the same amount of time also causes a dose dependent increase in 5-HT2a binding in cortex,(39) suggesting that this effect is mediated by GR, since dexamethasone is a potent GR agonist. Interestingly, the effect of corticosteroids on 5-HT2a receptor number seems to be specific for the cortex. Dexamethasone did not alter 5-HT2a binding in rat hippocampus. Given the relatively equal abundance of MR and GR in the hippocampus and the preponderance of GR in rat frontal cortex, it is plausible that 5-HT receptor regulation in the cortex is mediated through different mechanisms than in the hippocampus.
Stress, Serotonin and the LHPA Axis
In reviewing the clinical, psychological and biological literature on depressive illness, one factor that emerges as being closely associated with depression is stress. Stress and depression have been linked in a variety of ways. For example, both physical and psychological stressors have been shown to be temporally (and, perhaps, causally) related to the onset of depressive episodes.(61) Some studies have suggested that, at least for recurrent depression, stressful life events are more common in "non-endogenous depression." (23) Other studies have found that stressful life events are significantly correlated even with the first episode of psychotic/endogenous depression.(8) This is not to say that stress "causes" depression in people. Rather, stress is very likely interacting with an endogenous genetic predisposition, such that, in some vulnerable individuals, a stressor can precipitate a mood disorder (i.e., vulnerability + stress = depression). In fact, studies in twins by Kendler et al have demonstrated a clear interaction between genetic substrate and a recent stressful life event in the precipitation of a depressive episode: the more the genetic "loading" for depression, the more likely that a stressful event will trigger an episode.(35) There are, of course, cases in which the genetic "loading" or predisposition is so high that an episode of depression can occur in the absence of any apparent precipitating factors.
Another important link between depression and stress is the fact that both the LHPA and 5-HT systems, in addition to being involved in the pathophysiology of depression, are, also, as we have discussed above, critical contributors to the neurobiology of stress.(49) Therefore, studying the neurobiology of stress by focusing on these two systems has given us important clues into the pathophysiology of affective illness, shed light on the actions of antidepressants and begun to reveal how stress and mood disorders are related.
We have used chronic unpredictable stress, a postulated animal model of depression,(34),(2) to investigate the parallel changes in 5-HT and LHPA related molecules and to study how chronic stress and circulating glucocorticoids influence the 5-HT receptor system.(47) In this paradigm, rats are exposed to different mild to moderate stressors everyday, therefore making the stress "unpredictable" from day to day. Rats that undergo this treatment show LHPA overactivity, and increases in peripheral glucocorticoids, very similar to those found in MDD.(14),(2) We found that rats subjected to this paradigm show a significant decrease in 5-HT1a mRNA and binding in the hippocampus, as well as decreases in MR mRNA levels in this same region.
Antidepressant Medications, the LHPA Axis and 5-HT Receptors
The LHPA overactivity observed with chronic unpredictable stress can also be prevented by the chronic administration of either imipramine or desipramine, two tricyclic antidepressants.(45) Both desipramine and imipramine also reverse the stress induced downregulation of 5-HT1a in hippocampus, and the 5-HT2a upregulation in cortex. On the other hand, zimelidine and fluoxetine, two specific serotonin reuptake inhibitors, are unable to prevent the stress-induced elevation in corticosterone levels. This failure to prevent the LHPA overactivity is associated with a failure to restore the 5-HT receptor changes to baseline levels. This suggests that failure of an antidepressant to reverse the LHPA hypersecretion is associated with a failure to prevent the 5-HT receptor "dysregulation" secondary to chronic stress. We have proposed that this may be one of the neurobiological mechanisms underlying "treatment resistance" in patients with severe depression.(44)
There is some clinical evidence that these mechanisms may be operating in depressive disorders. Persistence of hypercortisolemia after antidepressant administration in depressed patients has been associated with relapse and poorer treatment outcome.(26),(64) In addition, some clinical studies have found that tricyclics are more effective than SSRIs in the treatment of melancholia.(17),(18),(65) Venlafaxine, an antidepressant with both norepinephrine and 5-HT reuptake activity, was reported to be more effective than fluoxetine in treating melancholic depression(15) and comorbid depression and anxiety.(67) Since melancholia and severity of depression are associated with a higher incidence of hypercortisolemia,(51) it is possible that the presence of a severely disturbed LHPA axis in this population may be contributing to the relative resistance to SSRI treatment. Interestingly, many augmentation strategies in treatment resistant patients are, in effect, attempts to broaden the biochemical profile of the pharmacological treatment, which may be more effective in reversing LHPA overactivity than a treatment whose main impact is in a single neurotransmitter system.
A Model of the Interplay Between Stress, the LHPA Axis, Serotonin and Antidepressants
Based on the animal and human studies reviewed here, we can generate a working model of the interplay between stress, the LHPA axis, 5-HT receptors and their potential interactions in suicide and depression. Figure 3 illustrates this model (solid arrows represent known effects, dashed arrows represent possible effects).
Figure 3.
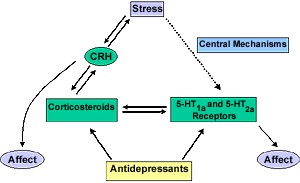
While it is possible that some monoamine receptor changes are due to antecedent changes in their endogenous ligands, we propose that many of the receptor changes observed may be a result of the LHPA overactivity present in at least some suicide victims, in particular those with a history of affective disorders. The rationale for this hypothesis derives from the above evidence, which can be summarized as follows:
- Depressed patients, as well as suicide victims, show evidence of overactivity of the LHPA axis.
- Chronic stress and/or high steroid levels in rats result in an alteration of specific 5-HT receptors (e.g., increases in cortical 5-HT2a, decreases in hippocampal 5-HT1a).
- Many human studies show the same receptor changes in the brain of suicide victims (increases in cortical 5-HT2a, decreases in hippocampal 5-HT1a) as found in hypercorticoid states.
- Chronic antidepressant administration causes opposite 5-HT receptor changes to those seen with chronic stress
- Antidepressant administration reverses the overactivity of the LHPA axis.
If indeed the 5-HT1a and 5-HT2a receptors have at least a partial role in controlling affective states (either directly or secondarily through other systems), then, their modulation by corticosteroids provides a potential mechanism by which these hormones may regulate mood. This, of course, does not exclude the possibility that stress can be simultaneously acting through other systems, such as the CRH receptors, thereby synergistically affecting mood and behavior. Antidepressants can counteract this phenomenon by affecting 5-HT receptor function directly and by simultaneously regulating stress induced corticosteroid secretion.
Conclusion
Corticosteroid modulation of 5-HT receptors has important implications for the pathophysiology and treatment of mood disorders and, perhaps, suicide. This may be one of the mechanisms by which stressful events can precipitate depressive episodes in some (genetically) vulnerable individuals and or precipitate suicidal behavior. Another implication is that altered 5-HT levels or metabolism do not necessarily have to be present for 5-HT receptor abnormalities to occur. Based on the animal data, it is apparent that specific 5-HT receptors may be directly regulated in response to alterations of corticosteroid levels. Thus, in depressed patients, normal levels of serotonin and its metabolites may not necessarily reflect normal central 5-HT activity.
An important therapeutic implication of this model is the prediction that agents, which can reduce the stress response and/or decrease LHPA activation, will be useful in the pharmacological treatment of anxiety, depression and, perhaps, suicidal behavior. In fact, patients with MDD, who are resistant to antidepressant treatment, have been reported to improve after receiving steroid suppression agents, like ketoconazole.(53),(71) However, these agents have many side effects and are often difficult for patients to tolerate. In this respect, CRH receptor antagonists, which are currently under development, may provide us with a new therapeutic weapon to treat these patients.(21),(13) These compounds could be used in conjunction with antidepressants, as adjuvants or augmenting agents, and may decrease treatment resistance. These agents may also be useful in monotherapy, since preventing hypercortisolemia may be translated into an improvement of monoaminergic receptor function. The use of modern biochemical and pharmacological tools, coupled with our increased understanding of the neurobiology of depression, should allow us to test these hypotheses, first in animal models and, then, directly in patients with affective illness.
The research reviewed in this paper was supported by a NARSAD Young Investigator Award, a Scientist Development Award (MH 01164) and by MH42251.